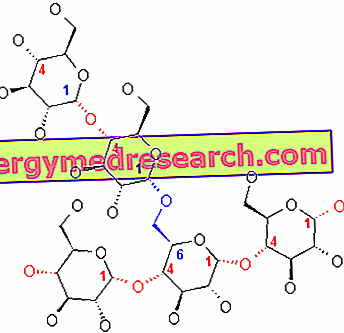
グリコーゲンは、主にα- 1,4グリコシド結合が存在し、α- 1,6グリコシド結合のために1:10の比で分枝しているα-グルコースの巨大分子(分子量約4億ダルトン)である。
グリコーゲンは予備物質を構成し、継続的に分解され再構成されます。 すべての体細胞量には、約100 gのグリコーゲンが含まれています。そのほとんどは移動可能な肝臓にあり、したがって他の臓器の予備として使用できます(筋肉のグリコーゲンは移動可能ではありません)。
グリコーゲンの分解と合成を触媒する酵素はすべて細胞質内にあるので、一方が活性である場合に一方の経路を不活性にする調節システムが必要です。グルコースが利用可能であれば、後者はグリコーゲンに変換されます。逆に、グルコースが必要な場合はグリコーゲンが分解されます(異化)。
グリコーゲンの分解に主に関与する酵素はグリコーゲンホスホリラーゼです。 この酵素は、溶解剤として無機オルトリン酸を用いてα- 1,4グリコシド結合を切断することができ、切断はリン酸分解を介して起こり、グルコース1-リン酸が得られる。
分岐点から5または6単位では、酵素グリコーゲンホスホリラーゼはもはや作用することができず、それゆえグリコーゲンから脱離し、トランスフェラーゼである枝切り酵素によって置き換えられる。この酵素の触媒部位には3つのサッカリド単位の最も近いグリコシド鎖への転移を可能にするヒスチジン(ヒスチジンはグルコース分子の最初の炭素を攻撃する)。 今述べた酵素はグリコシルトランスフェラーゼです。 この酵素の作用の終わりには、主鎖中のグルコースの6番目の炭素に結合している最初の炭素と共に、1つのグルコース単位のみが側鎖上に残る。 側鎖の最後のグルコース単位はα- 1,6グリコシダーゼ酵素の作用によって放出される(この酵素は枝切り酵素の第2部分である)。 グリコーゲン中の分枝が高分子の完全な分解から1:10の比にあるとすれば、約90%のグルコース1−リン酸と約10%のグルコースが得られる。
前述の酵素の作用により、グリコーゲン分子から側鎖を除去することが可能になる。 これらの酵素の活性は、鎖が完全に分解されるまで繰り返すことができます。
肝細胞について考えてみましょう。 それが細胞に入るとき、それはグルコース6-リン酸に変換され、そしてそれ故活性化される。 ホスホグルコムターゼの作用により、グルコース6-リン酸はグルコース1-リン酸に変換される。後者は生合成の非即時的前駆体である。 生合成では、活性化された形態の糖が使用され、それは二リン酸に結合した糖、通常はウリジル二リン酸(UDP)によって表される。 グルコース1-リン酸はその後UDP-グルコースに変換される。 この代謝産物は、成長しているグリコーゲンの非還元末端にUDP−グルコースを結合することができるグリコーゲンシンターゼの作用下にある:伸長グリコーゲンは、グルコシド単位およびUDPから得られる。 UDPは、酵素ヌクレオシドホスホキナーゼによってUTPに変換され、これが循環に戻る。
グリコーゲンの分解は、グルコース分子を放出してそれをグルコース1-リン酸に変換するグリコーゲンホスホリラーゼの作用によって起こる。 続いて、ホスホグルコムターゼはグルコース1−リン酸をグルコース6−リン酸に変換する。
グリコーゲンは、何よりも肝臓と筋肉の中で合成されます。体内には、筋肉量全体に分布した1〜1.2グラムのグリコーゲンがあります。
筋細胞のグリコーゲンはその細胞だけのエネルギーの蓄えを表し、肝臓に含まれるグリコーゲンは他の組織の蓄えでもあります。つまり、グルコースとして他の細胞に送ることができます。
筋肉でグリコーゲンの分解から得られたグルコース6リン酸は、エネルギー要件の場合には解糖に送られます。 肝臓では、グルコース6-リン酸は、 グルコース6-リン酸ホスファターゼ (肝細胞に特徴的な酵素)の作用によってグルコースに変換され、循環流に運ばれる。
グリコーゲンシンターゼとグリコーゲンホスホリラーゼの両方がグリコーゲンの非還元単位に作用するので、一方の経路の活性化と他方の経路の遮断を指令するホルモンシグナルが存在しなければならない(またはその逆)。
実験室では、グリコーゲンホスホリラーゼを利用し、非常に高濃度のグルコース1-リン酸を使用して、グリコーゲン鎖を長くすることが可能であった。
細胞内では、代謝産物の濃度が次の反応のバランスを右に(すなわちグリコーゲンの分解に向かって)シフトするようなものであるため、グリコーゲンホスホリラーゼは分解反応を触媒するだけである。

グリコーゲンホスホリラーゼの作用メカニズムを見てみましょう:アセタール酸素(グルコース単位間の橋渡しとして作用する)はホスホリル水素に結合します:反応中間体はカルボカチオンによって形成されます。ホスホリル(P 1)が非常に速く結合する末端)。
グリコーゲンホスホリラーゼはピリドキサールホスフェートである補因子を必要とする(この分子はトランスアミナーゼの補因子でもある):部分的にのみプロトン化されたホスホリルを有する(ピリドキサールホスフェートはそれに結合したプロトンの存在を正当化する疎水性環境に囲まれる) 。 このようなホスホリルはピリドキサールホスフェートの部分的にプロトン化されたホスホリルからのプロトンを取り戻すので、ホスホリル(Pi)はグリコーゲンにプロトンを生じさせることができる。 生理学的pHにおいて、ホスホリルがプロトンを失いそして完全に脱プロトン化されたままである可能性は非常に低い。
それでは、ホスホグルコムターゼの働きを見てみましょう。 この酵素は触媒部位にリン酸化セリン残基を有する。 セリンはグルコース1−リン酸(位置6)へのホスホリルを生じる:グルコース1,6−ビスホスフェートが短時間形成され、次いで位置1でホスホリルをとることによりセリンが再リン酸化される。 ホスホグルコースムターゼは両方向に作用することができる、すなわちグルコース1−リン酸をグルコース6−リン酸に変換すること、またはその逆である。 グルコース6-リン酸が生成されると、それは解糖に直接、筋肉に送られるか、または肝臓でグルコースに変換されます。
酵素ウリジルホスホグルコーストランスフェラーゼ(またはUDPグルコースピロホスホリラーゼ)は、ホスホリル攻撃によるグルコース1−リン酸のUTPへの転移反応を触媒する。
今述べた酵素はピロホスホリラーゼである:この名前は今述べたのと反対の反応がピロホスホリル化であるという事実による。
上記のようにして得られたUDPグルコースは、単糖単位のグリコーゲン鎖を長くすることができる。
ピロリン酸である生成物を排除することにより、UDPグルコースの形成に向けて反応を発展させることが可能である。 酵素ピロホスファターゼは、 ピロリン酸を2つのオルトリン酸分子(無水物の加水分解)に変換し、そうすることによって、ピロリン酸の濃度を非常に低く保ち、UDPグルコース形成プロセスが熱力学的に有利になる。
言及したように、UDPグルコースは、グリコーゲンシンターゼの作用により、グリコーゲン鎖を長くすることができる。
分岐点(1:10の比率で)は、グリコーゲン鎖が20〜25単位で構成されている場合、分岐酵素(触媒部位にヒスチジンを有する)が介在して一連の7を移動させることができるという事実による。 5〜6ユニットのさらに下流の−8グリコシドユニット:このようにして新たな分岐が生じる。
神経起源の理由のためにまたはエネルギーが肉体的な努力のために必要とされるならば、アドレナリンは副腎から分泌されます。
アドレナリン(およびノルエピネフリン)の標的細胞は、肝臓、筋肉および脂肪組織の細胞です(後者の場合、トリグリセリドの分解と脂肪酸の循環があります。ミトコンドリアでは、それゆえグルコースが生成されます6)。一方、脂肪細胞では、グルコース6-リン酸は、グルコース6-リン酸ホスファターゼ酵素の作用によってグルコースに変換され、組織に運ばれる。
それでは、アドレナリンの作用機序を見てみましょう。 アドレナリンは細胞膜(筋細胞および肝細胞)上に置かれた受容体に結合するようになり、これは細胞の外側から内側へのシグナルの翻訳を決定する。 プロテインキナーゼは活性化され、グリコーゲンの合成と分解を調節する系に同時に作用します。
グリコーゲンシンターゼは2つの形態で存在する:脱リン酸化型(活性)およびリン酸化型(不活性)。 プロテインキナーゼはグリコーゲンシンターゼをリン酸化しそしてその作用を遮断する。
グリコーゲンホスホリラーゼは、2つの形態、すなわちリン酸化セリンが存在する活性形態と、セリンが脱リン酸化される不活性形態とで存在し得る。 グリコーゲンホスホリラーゼは、酵素グリコーゲンホスホリラーゼキナーゼによって活性化され得る。 グリコーゲンホスホリラーゼキナーゼは、リン酸化されていれば活性であり、脱リン酸化されていれば不活性である。 プロテインキナーゼは基質としてグリコーゲンホスホリラーゼキナーゼを有する、すなわち後者はグリコーゲンホスホリラーゼを活性化することができ、そして後者を活性化することができる。
アドレナリンシグナルが終わったら、それが細胞内で持っている効果も終わらなければなりません:ホスファターゼ酵素が介入して、そしてタンパク質種に。